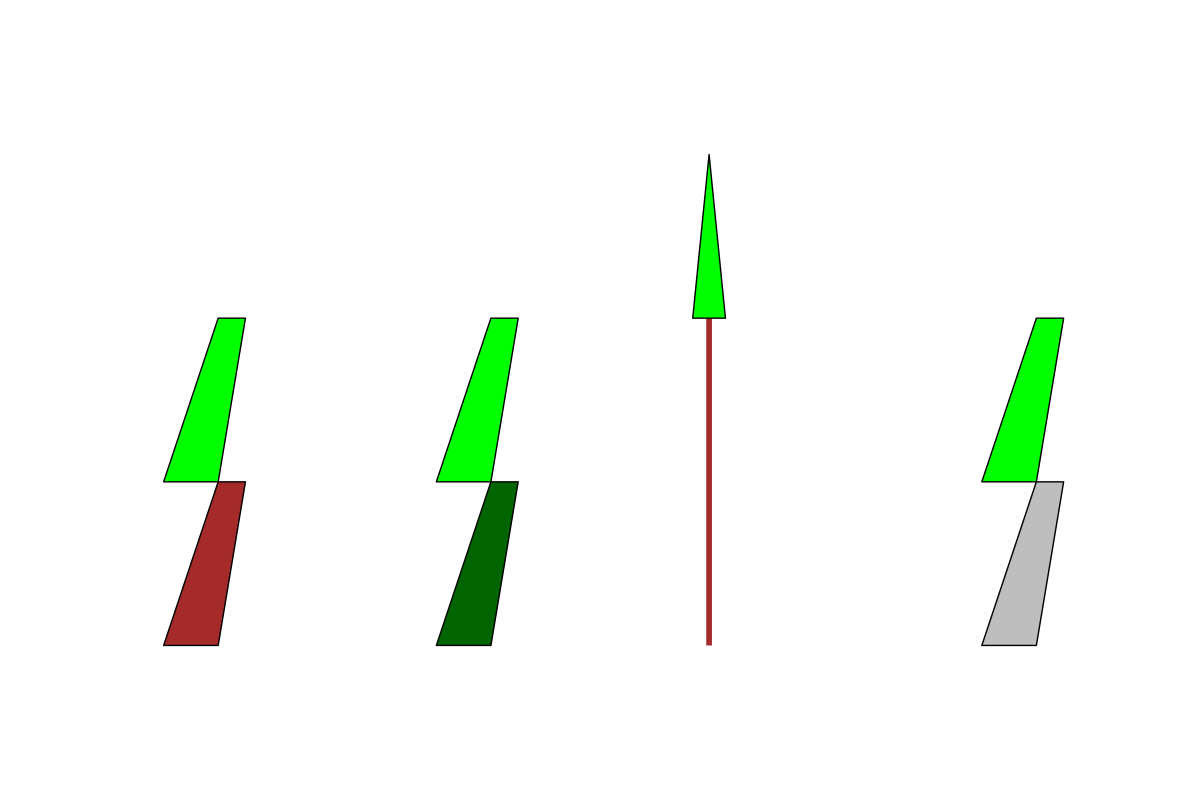I’m enjoying Robbie Phillips’ (and everyone else’s) grade rants on Instagram at the moment, and it’s got me thinking about some of the similarities between climbing grades and taxonomy – the science of biological classification.
Whereas climbing has numerous grading systems depending on what you’re climbing and where in the world you’re climbing it, biology has one. Based upon a system popularised by Carl Linnaeus in seventeen-hundred-and-something, organisms are named according to levels of increasing inclusivity. Species are at the finest end of the scale, with closely related species being included in the same genus; this is the basis of the ‘latin’ name that we’re all used to seeing. As an example, the lichen genus (pl. genera) Parmelia contains several related species: Parmelia sulcata, Parmelia saxatilis, Parmelia omphalodes etc. Related genera are grouped together into Families, Families into Orders, Orders into Classes, Classes into Phyla, and Phyla into Kingdoms. Understanding how organisms are organised within these different ranks is a key part of how we characterise biological diversity. It’s when we try to decide where any given organism fits within this classification system that the similarities to climbing grades become apparent
Both grades and taxonomy can be, to a greater or lesser extent, subjective. Whether something fits into one level or another only really makes sense within the context of whatever classification system it is that you’re using, and where the boundaries fall will depend entirely on the rules that you use. 7b+ or 7c if you’re short. E9 if you go one way, E7 if you go the other. HS if you can jam, E3 if you can’t. E1? HVS with pads mate. While the aim is to reach some sort of consensus grade there will always be folks who disagree, and defining species can be just as controversial. Although the levels of biological classification are more-or-less fixed, where the edges between them actually lie is up for debate and can be influenced by how you go about defining a species (what’s called a species concept). Do you define species by whether they interbreed (biological species concept), have similar DNA (phylogenetic species concept), look similar (morphological species concept), or live in similar places (ecological species concept)? In some tidy situations species might be well defined by all of these criteria, but often the answer changes depending on how you approach the question.
Me on Non-Metallic Silver at Irish Man’s Wall. E2 in the old guide, E1 in the new one although it was definitely E2 when I did it because it was my birthday.
Classification systems tend to evolve over time and just as climbing grades vary from one edition of the guide to another, so species names change. Lichen taxonomy in particular has undergone extensive revision in recent years. Lobaria virens has turned into Ricasolia virens (main picture). Alectoria nigricans became Gowardia nigricans, only to revert to its original name a few years later. The entire family Teloschistaceaewas broken up leading to new names for a remarkable 31 genera, the validity of which are still to be determined.
Alectoria (not Gowardia) nigricans
Like in climbing, debates about taxonomy can become heated. Just as a first ascensionist might not be happy about their route being downgraded, so a taxonomist could find themselves having to defend their new species circumscription against suggestions that it might be wrong. Once a new grade or revised species is finally settled on, the outcome can have repercussions for anyone using those classification systems. It can be demoralising to find that the route you’ve proudly been citing as the hardest thing you’ve ever done has been downgraded, and nothing frustrates a lichenologist like being told that you shouldn’t refer to your favourite lichen by the name you’ve been using quite happily for the past twenty-five years. Both grades and taxonomy can be cruel.
It’s all, of course, meaningless. A lichen doesn’t know what species it is any more than a rock knows how hard it is to climb, their names and grades imposed on them by us as a means of better interpreting our world. All part of this weird human desire to classify things. Yet meaningless as it is, I do wonder whether there’s some deep rooted evolutionary reason for this sort of behaviour. Non-human animals have been classifying things as food or not-food for hundreds of millions of years, and an innate desire to put things in boxes might literally be written in our DNA. Or it might not. At the end of the day, I just enjoy climbing rocks and looking at lichens. The rest is just academic.
I’ve recently been convinced to start using ChatGPT a bit more. I’ll confess, I don’t really ‘get’ A.I. yet. The actual nuts and bolts of the computational stuff involved are way over my head. I’m sure that it’ll revolutionise the world, but what about my own work? Despite some initial reservations I’m beginning to think there’s scope for using A.I. as a tool, but maybe not in the ways that I would have expected.
Unusually for a dry Saturday I’m not out climbing or lichen hunting, but sat at home nursing some lingering new year flu symptoms. In the interests of convalescing productively I thought I’d get ChatGPT to draw some lichens for me in R by asking the following question:
“Write me some code that will draw me a picture of the lichens Cladonia pyxidata, Xanothoria parietina, Rhizocarpon geographicum, and Lichina pygmaea using ggplot2.”
Me
ChatGPT was perfectly willing to carry out my request, but did offer me a word of caution:
“Creating detailed representations of multiple lichens in a single ggplot2 plot is quite complex due to the intricate structures of lichens. However, I can provide a basic example that includes a simplified representation of each lichen: Cladonia pyxidata, Xanthoria parietina, Rhizocarpon geographicum, and Lichina pygmaea.”
ChatGPT 3.5
Fair enough. The code generated was the following:
# Install and load ggplot2 if not already installed
if (!requireNamespace("ggplot2", quietly = TRUE)) {
install.packages("ggplot2")
}
library(ggplot2)
# Function to draw simplified representations of multiple lichens
draw_lichen_community <- function() {
ggplot() +
# Cladonia pyxidata
geom_polygon(
aes(x = c(2, 4, 5, 6, 7, 8, 9), y = c(2, 3, 4, 3, 4, 3, 2)),
fill = "green", color = "black"
) +
geom_point(
aes(x = c(4, 5, 6, 7, 8), y = c(3, 4, 3, 4, 3)),
size = 3, color = "brown"
) +
# Xanthoria parietina
geom_polygon(
aes(x = c(11, 12, 13, 14, 13), y = c(4, 5, 6, 5, 4)),
fill = "orange", color = "black"
) +
geom_point(
aes(x = 13, y = 5),
size = 3, color = "yellow"
) +
# Rhizocarpon geographicum
geom_polygon(
aes(x = c(18, 19, 20, 21, 20), y = c(3, 4, 5, 4, 3)),
fill = "gray", color = "black"
) +
geom_point(
aes(x = c(19, 20, 21), y = c(4, 5, 4)),
size = 3, color = "orange"
) +
# Lichina pygmaea
geom_polygon(
aes(x = c(25, 26, 27, 26), y = c(2, 3, 2, 1)),
fill = "purple", color = "black"
) +
geom_point(
aes(x = 26, y = 2),
size = 3, color = "pink"
) +
# Set axis limits
coord_cartesian(xlim = c(0, 30), ylim = c(0, 7)) +
# Remove axis labels and ticks
theme_void()
}
# Call the function to draw a simplified representation of multiple lichens
draw_lichen_community()
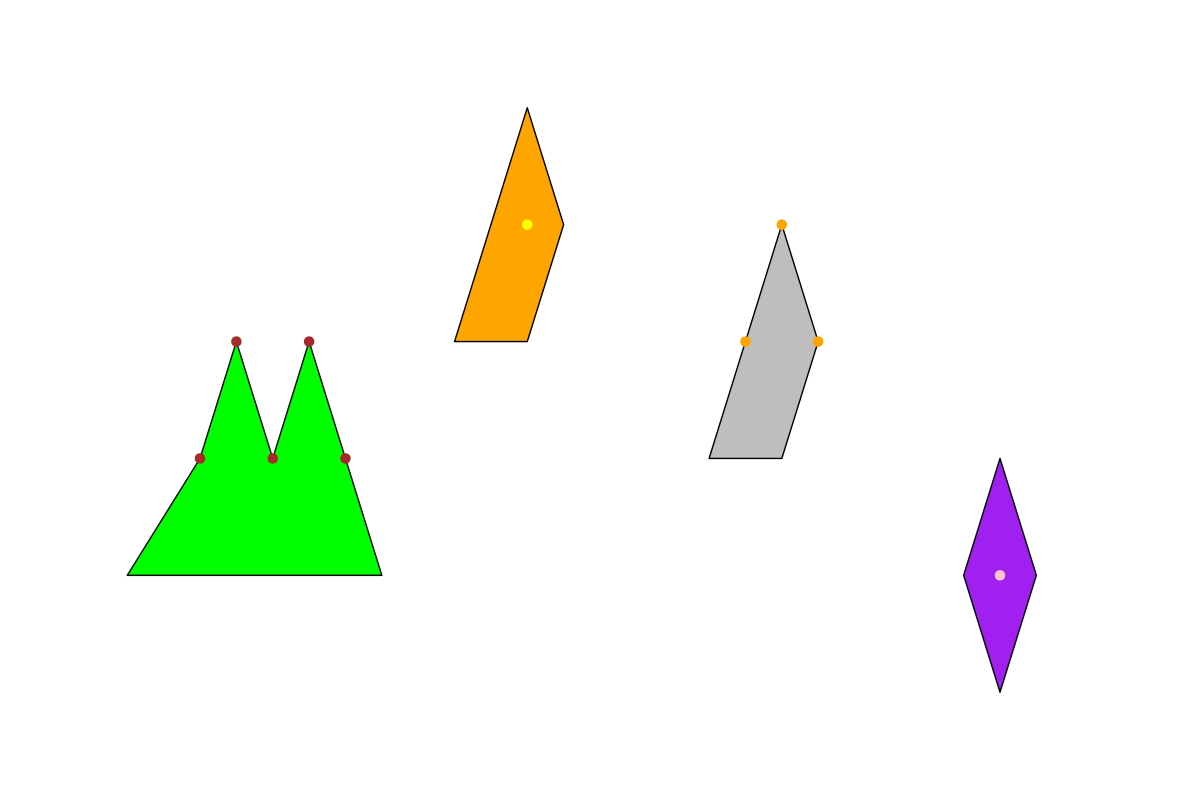
So far so good. I like the fact that it decided to refer to my plot as a lichen ‘community’; there’s obviously enough out there to train the A.I. that this is a word that’s commonly used when referring to assemblages of different species (although the habitat that supported a community containing all of these species together would be a rather odd one indeed). ChatGPT handed off with one final nugget:
“This code attempts to represent Cladonia pyxidata, Xanthoria parietina, Rhizocarpon geographicum, and Lichina pygmaea in a single plot. Adjustments and refinements may be necessary to better represent the specific features of each lichen. Keep in mind that this is a highly stylized and abstract representation for illustrative purposes.”
ChatGPT 3.5
This was the result:
A lichen community in ggplot2, as imagined by ChatGPT. Left to right: Cladonia pyxidata. Xanthoria parietina, Rhizocarpon geographicum, Lichina pygmaea.
On first glance this could appear either awful or amazing depending on your frame of mind, but there are some really interesting things here. First up is how colour has been chosen. The Cladonia is greenish and Xanthoria has come out in the expected orange. Grey-ish components of the Rhizocarpon geographicum thallus have been interpreted as dominant, while Lichina pygmaea is a fetching purple. Of course none of these colours are true to life, but as stylized choices I think they all work pretty well (with the exception of R. geographicum which I’ll come back to in a minute). There’s also been recognition that apothecia are different things to the thallus, being shown as different coloured points on top of the main thallus polygon. This has come out best on the Cladonia, with clear red/brown points contrasting with a green body, just like (well, sort of) the real thing. Xanthoria and Lichina are different shades of the thallus colour, which is fair. Interestingly, while the two-tone nature of Rhizocarpon has been recognised, it’s a bit muddled. While in real life the thallus is mostly yellow with black apothecia, this has been represented as the inverse in the plot, with a black (grey) thallus and yellow (orange) apothecia. While wrong, you can see why this could happen; R. geographicum thalli in real life are pretty variable and I think the uninitiated human would probably struggle to differentiate between which part was which. In its defence, ChatGPT isn’t a lichenologist and doesn’t really know what a thallus or an apothecium is anyway. The last thing to note is general morphology, which really hasn’t been depicted at all apart from in the ‘spikiness’ of the Cladonia which could be interpreted as multiple stalks.
The same lichens, but real. Left to right: Cladonia pyxidata. Xanthoria parietina, Rhizocarpon geographicum, Lichina pygmaea.
Some things to think about: ggplot2 is not the tool for drawing life-like representations of anything, let alone lichens, so I was already giving ChatGPT a pretty severe handicap. By doing so, it’s had an attempt at isolating some key features that it can use to answer the question. Interestingly, these are the key characteristics that we as people are drawn to as well; colour and shape. I often say to people that one of the reasons that lichens can be so hard for beginners is that there’s no archetypal lichen, no basic form that our minds jump to when we say the word ‘lichen’. Ask people what they think a lichen looks like and many will visualise Cladonia cups, while others think of a crust on a rock. There are no obvious straightforward rules that apply to all lichens as there are for something like trees (i.e. roots, trunk, branches, leaves). We might then use ChatGPT’s choices of what to put in these lichen plots as a way of identifying some rules of our own – that lichens can have two major parts, the thallus and the apothecia, and that lichens can come in lots of different colours.
As a test, to see how things went with what would, in my mind, be a simpler problem I thought I’d feed ChatGPT the same question but with some trees:
“Write me some code that will draw me a picture of the trees Quercus robur, Fraxinus excelsior, Pinus sylvestris, and Picea abies using ggplot2.”
Me
In its response ChatGPT automatically gave me back the common names of the species that I’d asked for, which was nice of it (I can see it struggling to do the same with lichens):
“Drawing realistic representations of trees using ggplot2 in R is quite challenging, as it’s primarily a tool for statistical and data visualization rather than detailed illustrations. However, I can provide a basic example that uses geometric shapes to represent simplified versions of oak (Quercus robur), ash (Fraxinus excelsior), pine (Pinus sylvestris), and spruce (Picea abies).”
ChatGPT 3.5
For anyone interested in the code, here it is:
# Install and load ggplot2 if not already installed
if (!requireNamespace("ggplot2", quietly = TRUE)) {
install.packages("ggplot2")
}
library(ggplot2)
# Function to draw simplified representations of multiple trees
draw_tree_community <- function() {
ggplot() +
# Quercus robur (Oak)
geom_rect(
aes(xmin = 2, xmax = 3, ymin = 1, ymax = 5),
fill = "brown", color = "black"
) +
# Fraxinus excelsior (Ash)
geom_rect(
aes(xmin = 7, xmax = 8, ymin = 1, ymax = 5),
fill = "gray", color = "black"
) +
# Pinus sylvestris (Pine)
geom_segment(
aes(x = 12, y = 1, xend = 12, yend = 5),
color = "green", size = 2
) +
geom_polygon(
aes(x = c(11, 12, 13), y = c(5, 8, 5)),
fill = "green", color = "black"
) +
# Picea abies (Spruce)
geom_segment(
aes(x = 17, y = 1, xend = 17, yend = 5),
color = "darkgreen", size = 2
) +
geom_polygon(
aes(x = c(16, 17, 18), y = c(5, 8, 5)),
fill = "darkgreen", color = "black"
) +
# Set axis limits
coord_cartesian(xlim = c(0, 20), ylim = c(0, 10)) +
# Remove axis labels and ticks
theme_void()
}
# Call the function to draw a simplified representation of multiple trees
draw_tree_community()
Again, ChatGPT has its disclaimer:
“This code uses rectangles and segments to represent simplified versions of Quercus robur, Fraxinus excelsior, Pinus sylvestris, and Picea abies. Adjustments and refinements may be necessary for a more accurate representation or to capture specific features of each tree. Keep in mind that this is a highly stylized and abstract representation for illustrative purposes. For detailed botanical illustrations of trees, specialized graphic tools and techniques may be more appropriate.”
ChatGPT 3.5
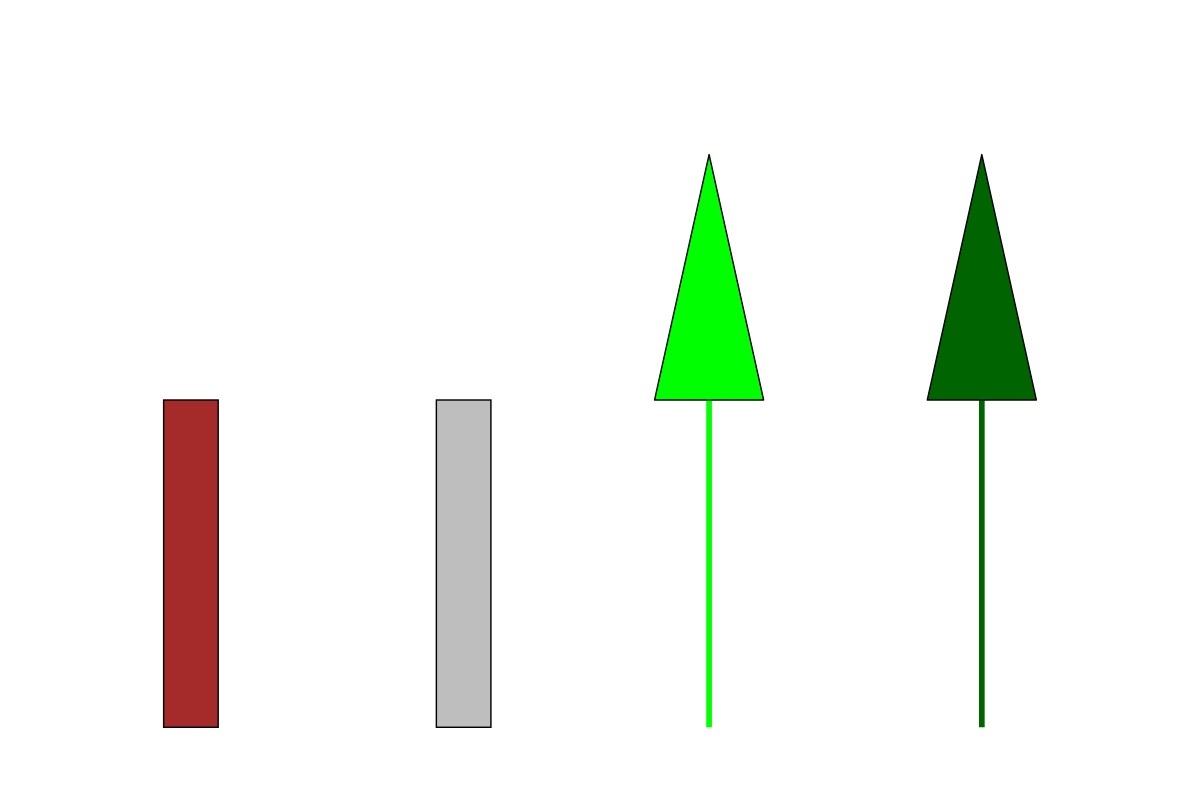
Some trees by ChatGPT in ggplot2. L-R: oak, ash, pine, spruce
Stylized is definitely the word. I was hoping that that it would draw the broadleaf trees with a circle on top and the conifers as a triangle, and while the latter part of this happened, both the oak and the ash have just been shown as trunks. It seems that ChatGPT isn’t really that interested in an archetypal anything but is just making it up as it goes along. I tried again, this time specifying that it show trunks and leaves:
the same trees with trunks and leaves. L-R: oak, ash, pine, spruce
Better, kind of, but still not what quite what I was expecting. I wasn’t sure how much I believed that ChatGPT really knew what it was doing here, so I asked the exact same question again but switched up the order of the trees to oak, spruce, ash, pine. This is what it came up with:
the same trees with trunks and leaves. L-R: oak, spruce, pine, ash
My confidence that ChatGPT was using any comprehensible logic to inform these plots at all was dwindling rapidly.
Whatever, in all of these cases we’ve gone in with a priori knowledge about what we’re looking at, which has really affected our interpretation (indeed, over-interpretation) of what ChatGPT has done . If someone showed you the lichen picture without telling you it was lichen, would you make that link? Probably not. The Cladonia could just as easily be a collapsed Christmas tree, or a bunch of basketballs on a grassy mountainside. Would the tree pictures fair any better? Perhaps, particularly with the addition of the ‘trunk and branches’ rule, but these are still highly abstract views. ChatGPT isn’t really telling us how to draw trees and lichens, but instead supplying an amalgamation of information about weirdly filtered copies of copies of something that somebody said was a tree or a lichen over infinite iterations, through the lens of 1,000 different pieces of code that were written to do something else entirely…. you get the idea.
Finally, I asked ChatGPT to draw me a lichen and a tree. ChatGPT thought I asked for a lichen on a tree and the outcome, a rusty rectangular Sputnik sporting a single tiny green orb, speaks for itself.
A lichen on a tree, apparently.
So what to conclude from all this? ChatGPT is good for many things, but it isn’t good at making things up. It needs prompts, both from you and from within the source material it’s drawing from in order to generate content. The less information it has on either side the worse it’s going to do, but at the same time it doesn’t know what to do with too much information. It’s also not particularly consistent; rules that might seem obvious to us aren’t automatically applied, and even if we supply a set of rules to follow, they likely won’t be adhered to in ways we expect. Arguably, the most useful thing that ChatGPT has done in this instance is to have made me write this blog and think about things in a way I wouldn’t have done otherwise. All scientific and artistic innovation throughout history has come about as a result of both the advances and limitations of the technology available at the time, with many interesting questions arising at the boundaries of what is possible. I think a particularly useful thing that can come out of AI is not specific answers to questions, but insight into how we actually frame those questions ourselves.
It’s Christmas, and I’ve been visiting my Mum in the village I grew up in. There’s a ‘new’ housing estate up the road – I say new, it was built over thirty years ago – with a wall at the entrance that’s got a really solid population of a crustose lichen called Ochrolechia parella (Fig.1). I’ve always quite liked this species, it’s fairly easy to recognise and a familiar sight when I’m out climbing on sea cliffs. While Christmas lunch was cooking, I thought I’d take the opportunity to try out a bit of lichenometry.
Fig.1. A wall, Hampshire
Lichenometry is the technique of using lichen growth rates as a way of measuring time. Lichens, as you may know, grow slowly. There’s a bunch of reasons for this (as well as a lot of exceptions) that I won’t go into here, but generally if you’re looking at a crustose lichen on a bit of rock you can assume it’s been there for a fairly long time. How long? The easiest way to get an idea of this is to go to a churchyard and look at gravestones. Older gravestones have more lichens on them. A few of those lichens might be pretty big and assuming that they’ve been there the longest we can get a good idea of their age by looking at the date on the stone, and from this you can work out their growth rate. This is the essence of lichenometry. It’s a technique that’s found a home in geography, particularly in dating the exposure of glacial moraine; if you have an estimated growth rate for a particular lichen that appears on the rocks of the moraine (geographers typically use Rhizocarpon geographicum, the map lichen) then you can have a stab at the date that those rocks were first revealed, giving you a picture of the dynamics of the glacier that deposited them. Neat.
Cool as this is, trying to work out dates from a single population of lichens is kind of impossible. In an uncomplicated world, a rock would be colonised by the first fungal spores on the day it was first exposed to the environment. All would germinate simultaneously and the lichens would grow at identical rates. No more lichens would establish themselves after that first cohort and nothing would interrupt their expansion. Every lichen would be a perfect circle and determining the age of that initial exposure would be straightforward.
This is of course wishful thinking (thankfully so – a world built on such simple rules would be missing all of the complexity that makes geography, ecology, and evolution such fascinating fields of study) and there are a bunch of things to confuse the situation. There was probably a lag prior to lichens getting established and while some individuals might have got off to an early start, others may have taken their time. Opportunities for colonisation don’t stop after the arrival of the first pioneers, and colonisation rates might increase once the first lichens reach maturity and start reproducing themselves locally. Once they’re established lichens might encounter environmental barriers, mosses, other lichens of both the same and different species, and the developing shapes of lichens are highly heterogeneous. With so much noise inherent in the technique, proper lichenometry studies use 1,000+ samples from multiple locations to calibrate measurements, and even then its been called into question how accurate they can actually be (Winchester, 2023).
But just because things aren’t perfect doesn’t mean that we shouldn’t see what we can find out, and I’ve always been a firm believer that useful insight can be obtained from even the crappiest of data. With that in mind, I went to measure some lichens. There’s not a lot in the way of moraine in Hampshire, but there is that wall at the edge of the housing estate.
My methods were simple:
Measure the diameter of O. parella thalli (plural of thallus, the word we use to refer to the body of a lichen) on the top of a waist-height brick wall in the south of England (Fig. 2).
When large thalli were clearly confluent (multiple lichens joined together), each part was measured as a separate thallus.
Only lichens that were easily identifiable as O. parella due to the presence of thick-rimmed apothecia (fruiting bodies) were counted.
Growth rates were calculated as the radius (i.e. half the diameter) in mm divided by the approximate length of time the wall had been standing (32 years) to give a growth rate in mm per year.
Eat Christmas lunch.
Fig. 2. Clockwise from top left: The wall again (TL), a small thallus (TR), a big thallus (BR), and a confluent thallus (BL).
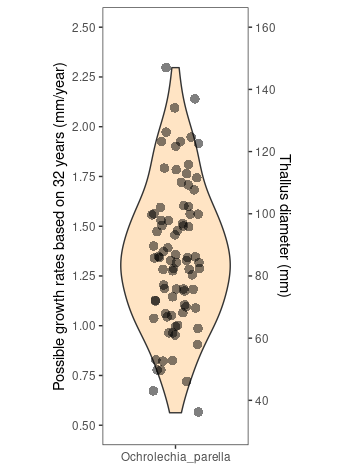
After filling up on cheese and mince pies I imported the measurements for 84 thalli into R and did some basic analysis. Lets pretend for a second that we live in that fantasy world where everything landed on the wall at the same time. Based on the size of the thalli and a known elapsed time of 32 years, if everything that’s there now had started growing simultaneously then we’d be looking at growth rates of O. parella ranging from 0.6 mm per year for the smallest thalli to 2.3 mm per year for the largest thalli (Fig. 3).
Fig. 3. A weird violin plot with some pretty unlikely growth rates
There are a couple of things to note here. First of all, this would mean the biggest thalli were growing four times faster than the smallest thalli. Is it likely that there’s this much variation in growth rate between individuals of the same species, in the same habitat? I guess it’s possible but I’d be inclined to think that this is probably supporting colonisation as a continuous process, with the smaller lichens having started growing later. Using the predicted growth rate of the biggest (and supposedly oldest) lichens of 2.3 mm per year, we can work out that an O. parella thallus 40 mm across might have been growing for somewhere between 8.5 and 9 years. This leads onto my second point; although these rates are slow I don’t think they’re CRAZY slow. Some lichens like R. geographicum, have average growth rates around 0.6 mm per year and as slow as 0.1mm per year in montane environments (Bradwell and Armstrong, 2007), so these measurements suggest O. parella is relatively rapid grower in the crusty lichen world. This fits with its habitat – rather than the extremely nutrient poor and snow covered environment of the mountains, the temperate south of England is much more favourable to faster rates of growth.
That’s not all we can learn from the frequency/size relationships of these lichens. Most thalli I measured were of a middling size; there were few very big lichens but also few small lichens. This is a bit different from most of the more thorough lichenometry studies that I’ve looked at, which tend to show a mean thallus diameter towards the smaller end of the spectrum. Have a look at these from lichens on bridges in Glen Coe (Fig. 4).
Fig. 4. A figure from Innes (1983) showing size frequency distributions of lichens on bridges in Glen Coe (N.B. this is a forty year old paper, I wonder what these lichens look like now?).
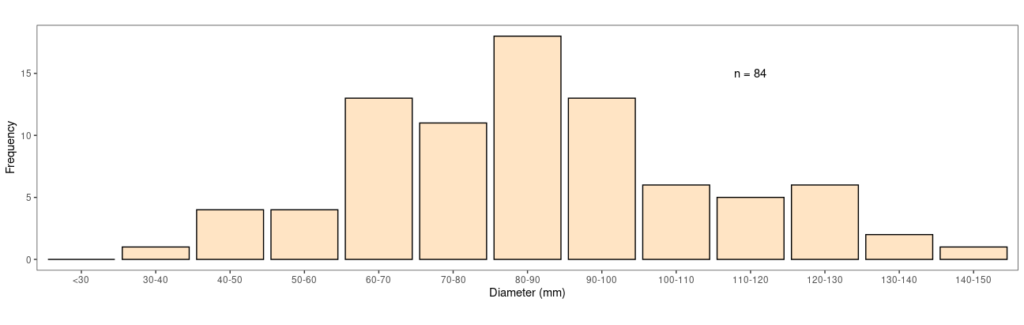
See how the graphs tend to lean to the left (counterintuitively we call this ‘right skewed’ data because most of the tail of the distribution is on the right)? I think this makes sense; you’re not going to end up with high numbers of very big individual lichens – there’s just not enough space – whereas you can pack quite a lot of small lichens into the same area. At early stages of colonisation the ‘elders’ of the lichen pack have little in the way of competition and are free to get big. Further down the line though, as more and more lichens get established and competition for space grows, individuals become restricted in how big they can get and the graph becomes biased towards higher frequencies of small thalli. So where are all the small thalli in my study? And why are there no lichens less than 30mm across (Fig. 5)?
Fig. 5. Size frequency distributions of lichens on the wall
One possibility is that the whole lichen community on the wall isn’t yet fully developed and that in time more smaller thalli will appear. This fits with the theory but still doesn’t explain the total absence of very small thalli, and I reckon there are some demographic factors that I haven’t taken into account. If you remember, I restricted my sampling to thalli with obvious fruiting bodies, ignoring anything that was infertile. There were lots of smaller white crusts and I have a suspicion that they are young O. parella thalli and it’s these that are missing from my distribution, which leads to one more piece of insight. Based on our calculations above, and assuming that our hypothesis is correct and the smallest thalli are infertile, we can work out that on this little wall O. parella is taking about 6.5 years to reach maturity. This is all quite speculative of course but not bad for twenty minutes staring at a wall on Christmas Day.
Bradwell T, Armstrong RA. 2007. Growth rates of Rhizocarpon geographicum lichens: a review with new data from Iceland. Journal of Quaternary Science22: 311–320.
Innes JL. 1983. Size Frequency Distributions as A Lichenometric Technique: An Assessment. Arctic and Alpine Research 15: 285–294.
Winchester V. 2023. Lichenometric Dating and Its Limitations and Problems: A Guide for Practitioners. Land 12: 611.
Since I started ‘doing lichens’ in 2020, the potential for combining lichenology and climbing has been at the front of my mind. The places that ropes let you access are seldom visited by lichenologists and I’ve always speculated that hidden gems might be revealed by a more ‘adventurous’ approach to lichen hunting. Earlier this summer, I was able to realise this potential on the basalt cliffs of Pentire Point.
Pentire is a cliff that’s on the radar of a lot of climbers in the South West and further afield, mainly due to the appeal of the Great Wall and its most famous routes: ‘Eroica’, ‘Black Magic’, and ‘Darkinbad the Brightdayler’. Andy Moles wrote about Darkinbad on UKC and as folks familiar with the work of James Joyce will know, anything that invokes Ulysses (or worse still Finnegan’s Wake) should be approached with considerable caution.
The Great Wall
My aspirations for the day were a bit more modest and I suggested to my pal Dexter that we climb a VS called ‘Our Stars, Our Sky’, a wandering a traverse above the sea. It was scorching hot and the north-facing sea level crags of Pentire offered a welcome escape from the sun. Dex led the zig-zagging first pitch which was covered in a slick of the ‘tar lichen’ Hydropunctaria maura (more on this in another blog) while I took the second, a fun traverse back above the rocks below.
Dex on Our Stars, Our Sky P1, some cool tension gashes in the rock between the ropes.Me on Our Stars, Our Sky P2
As we reached the top and scrambled back to our bags something caught my eye on a shady outcrop. I like to think I’m developing a pretty good eye for lichens and they tend to jump out at me from the background, especially if they’re something unusual. The form of this particular lichen was familiar but it seemed out of place. I was sure I was looking at Pectenia atlantica, a species I know pretty well from oceanic woodland in the northwest of Scotland.
Pectenia atlantica on a tree near Inverkirkaig, Assynt
I knew that this species could be found in the South West and I wrote off its rocky habit here as an interesting anomaly. Basalt’s an interesting rock – not as acidic as the granite that makes up so much of the climbable terrain in Cornwall – and where it’s found it often hosts unusual lichens, so seeing P. atlantica here wasn’t a completely ridiculous proposition. Something did feel different about this specimen but I couldn’t put a finger on it, again attributing its not-quite-right appearance to its growth on a not-quite-right substrate. I was pleased to go home with what was an unusual, if not groundbreaking, record.
Pectenia ligulata on coastal basalt at Pentire Head
Later that evening, I was reading through the Pannariaceae chapter of the Lichens of Great Britain and Ireland (a lichenologist’s bedtime reading is edgy stuff) and something jumped out at me: the description for Pectenia ligulata. Superficially similar to P. atlantica, P. ligulata is often found on coastal rocks, just like those that I’d found my specimen on. The main difference between the two is in some special structures that some lichens have; whereas P. atlantica has dense isidia, P. ligulata has spoon-shaped schizidia (sorry for the technical terms – lichenology has a lot of them and I’ll do some explainers in future blogs). I looked back at some photographs I’d taken, and my specimen did indeed appear to have spoon-shaped structures. Nice.
Get in close and you might be able to pick out little spoon-shaped scales all over the middle of the lichen.
However, before I settled on my new identification I had another problem to resolve; whereas there were several records for P. atlantica in the region, there were none for P. ligulata. In fact, the only record for the species in all of England was made nearly 25 years previously by Peter James on the Isles of Scilly.
Known UK distribution of Pectenia ligulata https://britishlichensociety.org.uk/resources/species-accounts/pectenia-ligulata
Faced with the prospect of such a rarity, I reached out to fellow lichenologists who might be familiar with the species. The agreement was unanimous – I did indeed have P. ligulata and its first record for mainland England. What a find! If there ever was justification for combining climbing and lichens this was it. I can’t wait to get out and explore more cliffs with a lichenologist’s eye; who knows what else might be out there.
Many thanks to Maxine Putnam, Heather Paul and Brian Coppins for their help with identifying the lichen, and to Dexter for continuing to climb with me despite my inclination to stare at rocks when I could be climbing.
A version of this post was published in the Winter 2023 edition of the British Lichen Society Bulletin
Hello! This is a blog about lichens. It’s also about climbing, geology, woodlands, mountains, ecology, sea cliffs, hillwalking, deep time, moors, running, mosses, rivers, boulders, insects… Welcome.
I wanted to start a blog for a few reasons:
I’ve been feeling that I’d like to write longer articles that don’t really work on Instagram or other short-form social media. I enjoy writing and want to practice it more.
I want to document some of my scientific research in a way that’s accessible, engaging, and useful to people outside of the academic bubble.
I’d like to build a permanent repository of ‘interesting stuff’ to act as a resource for me and for other people.
I’ve decided to call this blog The Vagrant Lichenologist. The Wikipedia page for vagrant lichen says this:
A vagrant lichen is a lichen that is either not attached to a substrate, or can become unattached then blow around, yet continue to grow and flourish. . . . Vagrant lichens generally occur in open and windswept habitats, all over the world
This is a pretty appropriate name because a) I can be kind of hard to pin down, b) I’m often found in open and windswept habitats, and c) I really, really like lichens.
There will be a lot about lichens here, but that’s not going to be the end of it. I’m also interested in how all things (including us) interact with the environment and each other. How communities, ecosystems, and landscapes grow and change over millennia. There are endless stories to be told about the planet and the things that make and inhabit it. Every aspect of our world is dependent on something else at every possible scale – the shape and form of a lichen adapted to an imperceptible microclimate or the structural and chemical properties of a rock that were determined half a billion years ago – and I find zooming in and out of these concepts super cool.
I hope this is fun. That’s the bottom line of why I’m writing this; the fact that I find learning about the natural world fun. I don’t like calling myself an expert (even in the things that I’m an expert in1) because I don’t think you need to be an expert to appreciate this stuff. I think it’s all really exciting, how everything works and fits together, and talking about exciting things is fun. The fact that I get to explore all of this when I’m out in the hills and climbing is the icing on the cake.
Finally, I would love this to become an interactive thing. Ask questions. Send me emails. Tell me about stuff I don’t know about. Tell me about your favourite rock. Ask me for a belay. Whatever.
See you out there x
I don’t have and formal geology training and I’ve only been working on lichens since 2020. Sure, I know a lot about how to assemble the genome of a cyanobacterium, or analyse communities of fungi based upon fragments of their DNA, but even in these fields I still have everything to learn. ↩︎